Date: March 1, 2024
Author: Roberto Barata
Last Update: August 5, 2024
How to cite: Barata, R. (2024). Understanding How Emotions Are Reinforced. Human-Animal Science.
Introduction
Contemporary trends in dog training suggest that emotions cannot be reinforced. This article will critically examine the traditional behaviorist approach alongside the contemporary neurocognitive perspective to elucidate the technical nuances of emotion reinforcement in dogs (and mammals in general), challenging the notion that emotions are impervious to such processes.
The traditional behaviorist framework posits that behaviors, rather than emotions, are the primary targets of reinforcement. According to this view, emotions are seen as byproducts of conditioning, secondary to observable behavioral changes. This perspective has historically guided dog training practices, focusing on modifying observable actions through reinforcement without directly considering the underlying emotional states.
Asserting that fear is not a behavior overlooks the broad definition of behavior in psychological terms, which includes both observable actions and internal states or responses. Fear, while often regarded as an emotional state, manifests through various behaviors, such as avoidance, freezing, or physiological responses, which can indeed be modified or influenced through conditioning processes.
As a different field, neuroscience offers a different perspective on understanding animal emotions by integrating insights from genetics, which reveal the hereditary aspects of emotional responses; neurobiology, which examines the brain structures and neural pathways involved in emotions; and cognitive science, which explores the processing and expression of emotions. It also suggests that emotions are not merely reactions to external stimuli but are also shaped by intrinsic factors such as genetic predispositions and cognitive evaluations. It posits that through understanding these complex interplays, reinforcement can indeed influence emotional states by altering the environmental and cognitive contexts in which emotional responses are elicited.
The adherence of some dog trainers to the belief that emotions cannot be reinforced may stem from several factors:
-
Methodological Tradition: A reliance on behaviorist principles prioritizing observable behavior changes over internal states, possibly due to behavior-based approaches’ direct measurability and simplicity.
-
Operational Challenges: The difficulty in quantifying and objectively assessing emotional changes in dogs, as opposed to clear, observable behaviors, makes it challenging to demonstrate emotional reinforcement empirically.
-
Conceptual Misunderstandings: There may be a lack of awareness, knowledge, study, or understanding of the neuroscience and neurocognitive research that illuminates how emotional states in dogs can be influenced by various forms of reinforcement, including social and environmental modifications.
-
Terminology Confusion and Subjective Interpretations: A significant barrier to the acceptance of emotional reinforcement among dog trainers is the conflation of terminology and reliance on subjective interpretations. Many trainers blend concepts from different fields without a clear distinction, leading to misunderstandings about what constitutes reinforcement, behavior, and emotion. The appeal to anecdotal evidence and compelling narratives in social media or popular publications further muddies the waters. Influential authors and online content creators, who echo similar beliefs about the non-reinforceability of emotions, often shape trainers’ perspectives more than the vast academic literature easily accessible to any proactive person. This reliance on less rigorous sources contributes to perpetuating misconceptions and incorporating inaccurate neuroscientific insights into training practices and discussions, which is the current problem in the field.
To clarify, while I possess a social and affective neuroscience background, it is not my primary area of expertise. My comfort level is limited to exploring neuroscience insofar as it is relevant to my practical work on animal behavior and training. I believe that we must concentrate on our respective areas of specialization and avoid overstepping our bounds, which has been a predominant tendency among dog trainers and dog training in general. My principal academic focus is ethology and human-animal studies, and my practical experience in animal training has been primarily with dog owners. My research aims to enhance my effectiveness and understanding in communicating with animals and people, as detailed in my book. This will be my sole article on the topic of emotions. Although I may update it as necessary, I do not intend to engage in further subjective or “ping-pong” discussions on this subject, especially in noisy environments such as social media.
I felt it was important to contribute to this subject by presenting the science found in academic literature rather than in dog training groups or courses. I will provide various perspectives, questions, challenges, references, and other resources to support your research and help you form an informed opinion. I will not present a definitive “truth,” as such an absolute does not exist. As discussed in this article, multiple theories and perspectives often exist on a single topic. Emotions are frequently discussed and sometimes exploited, with individuals even charging fees to share their insights—often without proper expertise. Reaching audiences can be challenging due to language and cultural differences, leading to misunderstandings and misinterpretations of terminology. Science does not offer absolute truths; therefore, trainers must step outside their comfort zones and critically evaluate their beliefs against credible sources rather than relying solely on social media groups or individuals.
Consulting and citing the relevant references individually is also necessary when challenging a viewpoint. Engaging in discussions about beliefs, addressing the messenger ad hominem, and dismissing the efforts to provide reliable information are counterproductive. This article aims to encourage a thoughtful and open-minded approach to these issues in the future. I hope you will engage with it proactively and be willing to assess the information presented critically.
Defining basic emotions
Charles Darwin was an early emotion researcher who explored whether basic emotions—fundamentally distinct and qualitatively different from one another—exist (fig.1). Basic emotions, if they exist, are categorized to simplify complex experiences, similar to how elements are classified in chemistry. However, it’s debated whether emotions fit this classification model. People tend to view emotions as separate categories, like happiness, sadness, and fear, possibly due to either inborn traits (as proposed by some researchers) or social construction (as suggested by Neimeyer, 1995). Researchers like Ekman (1994) and Panksepp (1994) have suggested criteria for identifying basic emotions, such as universality and distinct inborn expressions, though there is disagreement on which emotions qualify and how many there are.

Fig. 1. Darwin used photographs and drawings to illustrate the similarities between species in expressing emotions (anger/aggression). From Dalgleish (2004).
Defining basic emotions in humans is a complex challenge, and this complexity increases when applying these definitions to non-human animals. Emotions are internal, subjective experiences that are not directly observable. As a result, researchers must infer emotional states from observable behaviors, facial expressions, and physiological responses (Shepherd, 2017).
Basic human emotions are commonly categorized into discrete, universally recognized types: happiness, sadness, fear, anger, surprise, and disgust. However, there is an ongoing debate about whether these categories are truly universal and distinct or whether they are better described as continuous dimensions or blends of more fundamental emotional states (Reevy et al., 2010).
The challenge of defining emotions becomes even more pronounced when considering non-human animals. Although some species, such as dogs and cats, have been extensively studied and exhibit clear signs of emotional states akin to human emotions, others, like insects and fish, have more limited behavioral repertoires, making it less clear whether they experience emotions in a manner similar to humans (Adolphs, 2018).
Moreover, identifying behaviors associated with emotional states in non-human animals does not necessarily equate these behaviors with human emotions. For instance, a rat’s freezing response to a predator may indicate fear. Still, it could also be a reflexive response to danger that does not involve emotional experience in the same way as human fear.
The Challenge of Approaches from Different Fields
The growing interest in neuroscience has sparked interdisciplinary discussions involving psychology, sociology, philosophy, and ethology. This presents significant challenges, especially when professionals from non-neuroscientific fields engage with neuroscientific concepts without a deep understanding of the underlying principles and methodologies.
One major issue in interdisciplinary discussions is the risk of misinterpreting neuroscientific concepts. Neuroscience involves complex methodologies, including neuroimaging techniques (e.g., fMRI and PET scans), electrophysiological recordings, and molecular biology. Without knowledge of these methods, key concepts can be oversimplified or misrepresented. For instance, the term “neuroplasticity” is often misunderstood, leading to misconceptions about the brain’s capacity for change and recovery from neurological conditions (Kolb & Gibb, 2014).
Another concern is reductionism, where complex behaviors and mental processes are reduced to simple neural mechanisms. This reductionist view can undermine the multifaceted nature of human experience, which includes psychological, social, and environmental factors. Such oversimplification can result in deterministic views of human behavior, failing to account for the interplay between biological and environmental factors (Damasio, 1999).
Integrating neuroscience with fields such as law, education, and marketing introduces ethical concerns, particularly regarding the misuse or misrepresentation of neuroscientific findings. For example, neuroimaging used in legal contexts to assess a defendant’s mental state can raise ethical dilemmas about interpreting brain data ((Roskies & Morse, 2013). The potential for “neuroessentialism,” where individuals are judged solely based on neural characteristics, risks dehumanizing people by reducing them to biological substrates (Farah, 2011).
These brief examples show how effective communication across disciplines is necessary for successful collaboration. However, jargon and discipline-specific language can create barriers to understanding. When neuroscientific concepts are discussed without adequate explanation, it can lead to confusion and miscommunication, particularly in collaborative research efforts where shared terminology and methodology are fundamental (Sperber & Wilson, 1995).
Anthropomorphism in Neuroscience
The field of neuroscience has made significant strides in understanding brain function and behavior. However, integrating multiple theoretical frameworks, particularly when addressing phenomena like fear, can lead to confusion and oversimplification. This examination highlights the challenges of mixing theories and the impact of anthropomorphism, such as the metaphorical use of terms like “fidget.”
Anthropomorphism and anthropodenial are two interrelated but distinct concepts that shape how humans interpret the emotional lives of non-human entities. Though they represent opposing cognitive biases, both concepts are pivotal in influencing our understanding and treatment of animals.
Anthropomorphism involves attributing human-like characteristics, emotions, or behaviors to non-human animals. Anthropodenial, on the other hand, refers to the denial of human-like characteristics, emotions, or behaviors in non-human animals. This cognitive bias can obstruct the recognition and appreciation of the complexity and diversity of animal emotions.
If a term like “fidget” is used metaphorically to describe a neural mechanism with human-like characteristics, it may lead to oversimplified and misleading interpretations (Gordon, 2008). Such anthropomorphism can create a false sense of understanding and hinder accurate scientific discourse.
Mixing theories and using anthropomorphic language can result in oversimplification, which is problematic in neuroscience’s intricate domain. The brain’s complexity and many factors influencing emotional responses necessitate a nuanced approach. Oversimplification, whether through theory mixing or anthropomorphism, can obscure the multifaceted nature of emotional processing and limit clinical interventions (LeDoux, 2012).
Do All Emotions Need to Be Expressed Physically to Be Considered Emotions?
The question of whether emotions must be expressed physically to be considered valid emotions invites a nuanced exploration into the nature of emotions and their expressions. Emotions encompass subjective experiences, physiological responses, and behavioral expressions. The James-Lange theory posits that physiological arousal precedes and constitutes emotional experience, suggesting that bodily changes are crucial to feeling emotions (James, 1884). Conversely, the Cannon-Bard theory argues that emotional experiences and physiological responses occur independently and simultaneously (Cannon, 1927). These theories indicate that while physical responses often accompany emotional experiences, they are not strictly necessary for the existence of emotions.
The distinction between emotional experience and emotional expression is essential in understanding whether physical expression is required for emotions to be valid. Emotional experience pertains to internal, subjective feelings, whereas emotional expression involves outward manifestations such as facial expressions and body language (Ekman, 1992). Research indicates that individuals can experience various emotions internally, even without physical expression. For instance, individuals with neurological conditions, such as those affecting facial musculature, may find it challenging to express emotions outwardly but still experience them internally (Hennenlotter et al., 2009). This evidence supports the notion that emotional validity is not contingent upon physical expression alone.
Philosophical views also offer critical insights into this discussion. Existentialist philosopher Sartre (1943) argued that emotions are deeply personal and subjective, not fully captured by external expressions. According to Sartre, the essence of an emotion resides in the individual’s internal experience rather than its outward manifestation. Similarly, Merleau-Ponty (1962) emphasized that while physical expressions can enhance communication, they are not necessary for the existence of emotions. From these perspectives, the internal experience of emotions holds primacy over their external expression.
Neuroscientific research has identified specific neural circuits associated with emotional experiences, such as the amygdala’s role in processing emotions like fear and anger (LeDoux, 1996). Studies demonstrate that emotions can be experienced without physical expression, reinforcing the idea that emotions are not solely dependent on outward manifestations.
Sociological perspectives highlight that cultural norms and social contexts shape emotional expression. For example, cultural expectations can influence how and when emotions are outwardly displayed, indicating that physical expression is culturally contingent rather than universally required (Hochschild, 1983).
In clinical psychology, cognitive-behavioral therapy (CBT) emphasizes the importance of understanding and processing emotions internally, regardless of their physical expression (Beck, 2011). This approach supports the notion that emotions are valid and meaningful even without physical manifestation.
The interplay between sensory processing and emotional experience can also be useful in this discussion. Sensory processing involves the detection of stimuli through senses such as sight and touch, which are then interpreted by the brain to form emotional responses (Schacter & Singer, 1962). Physical sensations, such as warmth or texture, can evoke emotional responses and illustrate the connection between physical sensations and emotional states. Perception also shapes emotional experiences by interpreting sensory inputs based on past experiences and beliefs (Neisser, 1976). Damasio’s concept of somatic markers illustrates the interplay between physical sensations and emotional experiences. Somatic markers are bodily sensations associated with emotions that guide decision-making and influence emotional responses (Damasio, 1994). This interplay highlights the importance of understanding emotions as encompassing physiological and emotional dimensions, though it does not imply that physical expression is necessary for validity. Therefore, the assertion that all emotions must be expressed physically to be considered valid is a reductionist perspective that overlooks the complexity of emotional experiences.
Panksepp’s Theory in Dog Training
In recent discussions about dog training methodologies, I’ve observed a growing trend of positioning Jaak Panksepp’s theory of affective neuroscience as the sole acceptable framework for discussion. This trend has led to the rejection of alternative perspectives, even those from credible authors with differing views. During a brief exploration of social media, it became evident that proponents of other theories are often met with skepticism or outright rejection.
My review of various neuroscientific encyclopedias revealed that Panksepp’s work is not extensively referenced within the broader field of neuroscience (Della Sala, 2020; Ray, 2024; Reevy, Ozer, & Ito, 2010; Shepherd, 2017). This absence might reflect a more nuanced understanding of emotional processes that are not fully captured by any single theory. It makes sense because the study of emotions, such as fear, encompasses a range of perspectives. For instance, Adolphs (2013) highlights at least nine distinct views on fear, each contributing to a more comprehensive understanding of emotional responses.
Restricting our focus to a singular theory, such as Panksepp’s, poses risks akin to those observed in past debates. I often give the example of the discussion on the “theory of dominance” in dog training, which was once widely spread and debated with Mech’s work being always referred to, but later revealed to be a misinterpretation of Mech’s work, as he clearly confirmed that people misunderstood his writings (Barata, 2012). Panksepp’s work, too, is being similarly isolated and potentially misapplied. The current trend of treating Panksepp’s theory as the definitive approach overlooks valuable insights from other perspectives and limits our understanding of canine behavior, leading to more subjective assumptions and pushing us even further away from what we can observe and measure, which is already complex.
The term “affective neuroscience,” popularized by Jaak Panksepp, refers to the interdisciplinary study that integrates the examination of emotions (affect) with neuroscience to elucidate the neural mechanisms underlying emotional processes. Panksepp’s work in the early 1990s established this field, focusing on the emotional systems in animals and their neural correlates. His identification of seven primary emotional systems—SEEKING, FEAR, RAGE, LUST, CARE, PANIC/GRIEF, and PLAY—aimed to provide a foundational framework for understanding the neurobiological bases of behavior (Panksepp, 1998, 2005).
The study of emotions and their neural correlates has earlier theoretical foundations laid, for instance, by psychologists and neurologists such as William James and Carl Lange in the late 19th century, who proposed physiological bases for emotions. Subsequent advancements in the mid-20th century included James Papez’s formulation of the Papez circuit, which mapped a neural circuit involved in emotional expression, and Paul MacLean’s expansion of the limbic system concept. The research of the 1960s and 1970s further refined our understanding by exploring specific brain structures like the amygdala and their roles in emotional processing (LeDoux, 2012).
Despite Panksepp’s significant contributions, several limitations of his theory become apparent, particularly in the context of practical applications such as dog training. Firstly, Panksepp’s model tends to oversimplify the complexities of canine behavior by focusing on primary emotional systems, potentially neglecting the multifaceted interplay of cognitive processes, environmental influences, and learned behaviors. This simplification contrasts with cognitive learning theories, such as those proposed by Pavlov and Skinner, which emphasize the role of reinforcement, punishment, and associative learning in shaping behavior (Pavlov, 1927; Skinner, 1953).
Panksepp’s theory has also been critiqued for its insufficient consideration of behavioral ecology and ethology, which examine natural behaviors within ecological contexts and emphasize species-specific adaptations. Additionally, the theory’s failure to account for individual variation and breed-specific behaviors can lead to less effective training strategies, as dogs exhibit significant diversity due to genetic and selective breeding factors (Shettleworth, 2010).
Panksepp’s dualistic perspective, which separates mind and body, may further constrain the applicability of his theory. While historically significant, this dualistic approach may limit the acceptance of integrative models that combine cognitive, emotional, and contextual factors (Damasio, 1994; Rolls, 2013). Additionally, Panksepp’s research methodologies, which often involve invasive procedures, raise ethical concerns about the balance between scientific inquiry and the welfare of the animals studied (Panksepp & Biven, 2012).
Furthermore, Panksepp’s focus on subcortical emotional systems might overshadow the role of higher cognitive functions located in the neocortex. Cognitive processes such as reasoning, problem-solving, and memory play important roles in how dogs learn and interact with their environment (LeDoux, 2012; Barrett & Satpute, 2013). For example, claims that dogs lack cognitive coping mechanisms for stress simplify their complex behavioral and cognitive responses.
Contrary to this view, evidence shows that dogs possess sophisticated cognitive abilities that aid in managing stress. Research has revealed that dogs are capable of cognitive processing beyond mere instinctual reactions. Training provides further evidence of dogs’ cognitive abilities. Through training and conditioning, dogs learn complex tasks and adapt their behavior based on specific signals. This process involves memory and problem-solving skills. Miklosi (2007) demonstrates that dogs can engage in problem-solving, memory, and learning, which are crucial for handling stress. These cognitive skills imply that dogs use more than just basic responses to cope with stress. Dogs also exhibit significant behavioral flexibility in response to changing environments, indicating cognitive processing. They can adapt their behavior based on new experiences and information. For example, dogs can modify their actions in response to altered circumstances, like age, reflecting their ability to adjust their coping strategies (Wallis et al., 2013). This flexibility suggests that dogs employ cognitive mechanisms when dealing with stress.
Comparative research supports the view that domestic mammals have advanced cognitive functions. This research on the five sociocognitive abilities of domestic mammals shows that some species can recognize us or detect and interpret human emotions or signals (Jardat & Lansade, 2021). These findings challenge the idea that non-humans rely solely on instinctual responses. The emotional lives of dogs further illustrate their cognitive coping mechanisms. Dogs experience emotions such as stress and anxiety, which influence their coping strategies. A study of dogs’ emotional responses, including their capacity for empathy and stress management, supports the argument that cognitive processes are involved in their coping mechanisms (Bradshaw, 2011). Observations of dogs in natural settings provide additional insights into their stress-coping strategies. Dogs employ various coping behaviors, such as seeking comfort or changing their actions in response to stressors. These behaviors suggest that cognitive strategies affect how dogs handle stress and adapt to their environment.
As a final note, I want to mention that critical neuroscience examines the intersection of cognitive neuroscience with broader philosophical, cultural, and social contexts. It challenges the reductionist view that the mind can be fully explained by brain functions alone, advocating instead for a more holistic understanding that incorporates the dynamic interactions between neural processes, the body, and the environment. This perspective also critiques how neuroscience is sometimes employed ideologically to support or justify social and economic structures, such as flexible capitalism. Critical neuroscience seeks to expose how scientific “facts” are constructed and urges a more nuanced and reflective approach to understanding the mind and brain (Choudhury & Slaby, 2011).
What is “Reinforcement”?
Reinforcement is the process of strengthening or increasing a behavior through a stimulus. There is a tendency to associate reinforcement only with operant conditioning. Skinner did not introduce the concept, even though he defined it precisely. Hull, Watson, and Thorndike had already mentioned reinforcement as a stimulus that increases a response. Arguing against the perspective that reinforcement should be exclusively associated with operant conditioning requires a nuanced understanding of learning and behavior modification principles. At its core, reinforcement is about increasing the likelihood of a behavior or response through stimuli, a principle not limited to the deliberate manipulation of outcomes as seen in operant conditioning.
Furthermore, the argument that reinforcement should be an exclusive term for operant conditioning overlooks the adaptive significance of learning and physiological mechanisms. Both classical and operant conditioning have evolved to help organisms learn from their environment in ways that enhance survival and reproduction. In terms of mechanism and applicability, the concept of reinforcement is fundamental to understanding how organisms adapt to their environments. Therefore, limiting the concept of reinforcement to operant conditioning, as widely read in social media, is inaccurate and neglects the complexity and diversity of learning and behavior modification mechanisms.
Classical conditioning, a learning process first described by Ivan Pavlov, involves the association of a neutral stimulus with an unconditional stimulus to elicit a conditional response. In this framework, the unconditional stimulus (US) naturally triggers a response, while the conditional stimulus (CS) is initially neutral but becomes associated with the US through repeated pairings. For example, in Pavlov’s famous experiment, the sound of a bell (CS) was paired with the presentation of food (US), leading to the dogs salivating (conditional response, CR) at the sound of the bell alone.
Some researchers extend the concept of reinforcement to classical conditioning. In this context, the US can be viewed as a reinforcer because it strengthens the CS and CR association. When the US is presented following the CS, it reinforces the learning process by providing a meaningful outcome that the organism learns to anticipate. For instance, when the bell is rung before the food is presented, the food serves as a reinforcer that solidifies the connection between the bell and the salivation response. This reinforcement is necessary for establishing the conditional response, as it helps the organism learn that the CS predicts the arrival of the US. However, it is important to recognize that classical and operant conditioning terminology and concepts are distinct. Classical conditioning focuses on the association between stimuli, while operant conditioning emphasizes the role of consequences in shaping behavior. Using the term “reinforcer” in classical conditioning can lead to confusion, as it blurs the lines between these two fundamental learning processes.
Classical conditioning also involves the modification of behavior. The process of pairing a neutral stimulus with an unconditional stimulus to produce a conditional response can be seen as reinforcing because it increases the likelihood of the conditional response to the previously neutral stimulus (Olson, 2016). Moreover, the distinction between classical and operant conditioning is not always clear-cut in real-world learning scenarios. Many behaviors are influenced by a combination of associative learning and the consequences of actions, suggesting that reinforcement operates along a continuum rather than within strictly defined categories. For example, the development of phobias can be explained through classical conditioning, but the maintenance of avoidance behavior can be reinforced operantly by the relief it provides, demonstrating the interconnectedness of these learning processes.
The original Russian term used by Pavlov that corresponds to “reinforcer” is “подкрепление” (pronounced “podkreplenie”). This term is often translated as “reinforcement” in English. While Pavlov’s work laid the foundation for concepts related to reinforcement in behaviorism, he did not extensively develop these ideas in the same way other psychologists would later explore them. Instead, his focus was primarily on the mechanisms of classical conditioning and the associations formed between stimuli and responses. As it is understood today, the concept of reinforcement was not a primary focus of Pavlov’s terminology or theoretical framework. However, the term “подкрепление” is indeed associated with his research on conditional responses and learning processes.
Pavlov used the term to describe the strengthening of an already-learned but weakening response, not the modern definition of selecting and strengthening new behaviors. Although the widespread use of the term reinforcement in learning theory is traceable to Pavlov’s influence (1927), he was not himself a reinforcement theorist in the sense that came to be accepted in American psychology. He identified reinforcement with the occurrence of the unconditional stimulus and its elicited response. Key terms like “reinforced” and “reinforcement,” which now serve as the approved labels for the central concept in our discipline, arose largely by accident. They are universally, and in a sense, correctly attributed to Pavlov (1927/1960), but in the historical compendium of his addresses published in 1928, the word “reinforcement” appears only once and then as a rather loose translation of the original Russian text (Razran, 1955). According to Razran, Pavlov and his students he has never used the term “reinforcement” for the trials preceding the full-fledged emergence of the CR. In the more systematic series of lectures published in 1927, Pavlov often used the notations “reflex reinforced” or “stimulus reinforced” in the trial-by-trial protocols for individual experiments, but “reinforcement” never appeared as a theoretical concept. It was used to describe the practice of reinvigorating the conditional salivary secretion after it had been weakened by repeated elicitations by presenting one or more trials where the original unconditional stimulus followed the conditional stimulus. If the unconditional stimulus was not presented on a given trial, the effectiveness of the conditional stimulus declined, and the trial was classified as one of extinction, another term apparently borrowed directly from a common language.
Some argue that the acquisition of conditioned responding should be broken down into components, including the point of acquisition and the asymptotic response magnitude (Gallistel, Fairhurst, & Balsam, 2004). Generally, increasing US intensity or number tends to increase the magnitude of conditioned responding (Mackintosh, 1974, pp. 70-71). However, although Morris and Bouton (2006) detected effects of US magnitude on the point of acquisition in aversive and appetitive procedures with rats, the extent to which US magnitude influences the point of acquisition is still unclear. Unlike US intensity, the effects of increasing US duration are not consistent across procedures (Mackintosh, 1974).
Perhaps some of these terms are lost in translation or mixed with Tolochinov, Vurfson, Babkin, Boldyrev, or Kashereninova’s work (Windholz, 1989). For instance, many English-language textbooks use terms like unconditioned stimulus” and “unconditioned response.” These terms originated from translation errors when Pavlov’s original Russian terms (conditional, not conditioned) were translated into English (Fitzpatrick, 1990; Goldman, 2012). Reinforcement is a term that can mean many things in both its technical and nontechnical usage. It always involves something related to strengthening, but even in lay language, it can legitimately refer to the act of strengthening, the state of being strengthened, or the thing that strengthens. In its technical use in psychology, reinforcement has been employed in all these senses and many other specialized ones. For example, in the dictionary of psychological terms compiled by English and English (1958), two and a half pages are required to list and describe the various usages. They cover nine psychological meanings of the term “reinforcement” and 32 separate entries of its various combined forms, such as aperiodic, primary, and serial reinforcement. They prefer the usage based on the Pavlovian paradigm: “the natural occurrence or the experimental presentation of the unconditional stimulus along with the conditional stimulus; or the strengthening of the conditional response relationship thereby.” For the remaining special meanings with enough operational specificity to be useful, English and English recommend using alternate words such as facilitation, reward, drive or tension reduction, confirming reaction, and goal attainment.
You can read more about it in the following references: Dinsmoor (2004), Pavlov (1960), Rescorla & Wagner (1972), Schoenfeld (1978), Schoenfeld (1995), Windholz (1989), and Yokoyama (2023).
Mechanisms of Reinforcement
As mentioned above, the term “reinforcer” or “reinforcement” generally refers to something that enhances or strengthens a behavior. However, the mechanisms of reinforcement vary across different disciplines. Although the core idea of reinforcement as a means to strengthen behavior remains consistent, the interpretations and mechanisms can vary across disciplines due to their distinct focuses. Each field may define and interpret these mechanisms differently, focusing on distinct processes or principles that underpin how reinforcement operates within that domain. Therefore, the specific mechanisms of how each field’s particular theoretical and empirical frameworks shape reinforcement functions.
Neuroscientific Perspective
In neuroscience, reinforcement is closely linked to the brain’s reward circuitry, which involves several key structures, including the nucleus accumbens, ventral tegmental area (VTA), and prefrontal cortex. “Reward circuitry” refers to the network of brain structures and pathways involved in processing rewards, motivating behaviors, and reinforcing learning. This circuitry is vital for survival, as it helps organisms seek out and repeat beneficial behaviors, such as eating, mating, and social interactions. The reward circuitry is heavily implicated in the experience of pleasure, motivation, and reinforcement learning. Mechanisms include:
-
Dopaminergic Pathways: The release of dopamine in response to rewarding stimuli is a fundamental reinforcement mechanism. Dopamine signals the perceived value of a reward and reinforces behaviors that lead to its attainment.
-
Neuroplasticity: Reinforcement can change synaptic strength and neural circuitry, facilitating learning and memory. Long-term potentiation (LTP) enhances synaptic transmission and is associated with the reinforcement of learned behaviors.
-
Conditional Responses: Classical conditioning studies, such as those conducted by Pavlov, demonstrate how neural pathways can be reinforced through associations between stimuli and reinforcers, leading to conditional responses.
Neural Mechanisms of Reinforcement
The brain regions involved in reinforcement processes are part of the reward circuitry:
-
Ventral Tegmental Area (VTA): The VTA contains dopamine-producing neurons that project to various brain regions in the midbrain. It is a primary source of dopamine, a neurotransmitter central to reward processing.
-
Nucleus Accumbens: This basal ganglia area is crucial for processing rewards, pleasure, and reinforcement learning. It receives dopamine signals from the VTA.
-
Prefrontal Cortex (PFC): This area, specifically the orbitofrontal cortex (OFC), is involved in decision-making, evaluating rewards, and planning. It integrates information about reward value and helps regulate behavior based on expected outcomes.
-
Amygdala: This structure is involved in emotional processing and associating stimuli with reinforcers or punishments, playing a role in both reinforcement and fear conditioning.
-
Hippocampus: Important for memory formation and spatial navigation, the hippocampus interacts with the reward circuitry to help form memories of rewarding experiences, guiding future behavior.
-
Striatum: This includes the caudate nucleus and putamen, which are involved in motor control and reinforcement-based learning. The striatum integrates reward information to influence movement and behavior.
-
Hypothalamus: Involved in regulating homeostatic functions like hunger and thirst, the hypothalamus interacts with the reward circuitry to influence motivated behaviors related to survival.
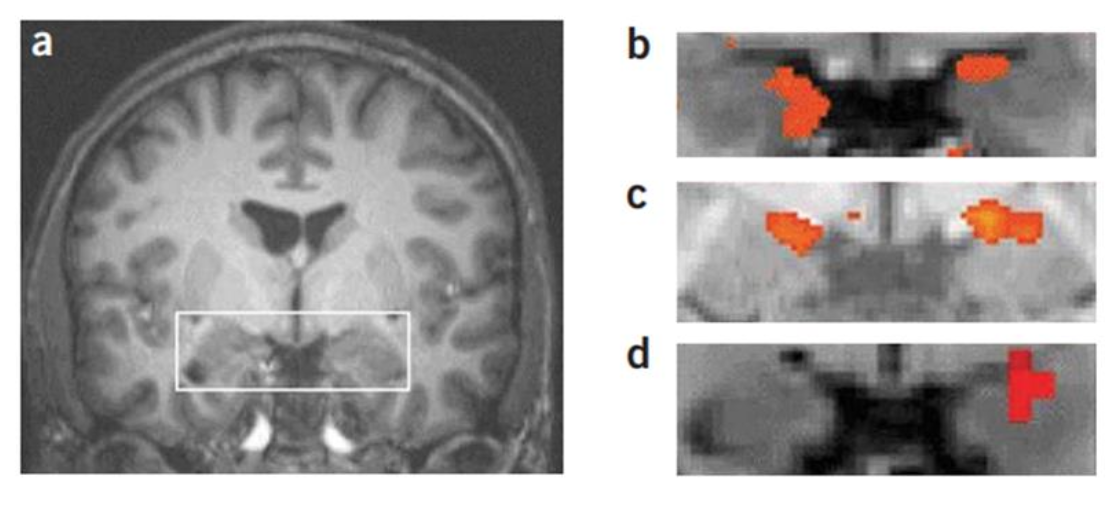
Fig. 2. Fear learning in the human amygdala. A) The medial temporal lobe, including the amygdala, is bilateral. B-D) Activation of the amygdala towards the CS is observed bilaterally after fear conditioning (b) and observational fear learning (c) and unilaterally (d) in the left amygdala after instructed fear. From Olson et al. (2007).
Limbic System Involvement
-
The limbic system, a set of brain structures including the amygdala, hippocampus, and prefrontal cortex, plays an important role in emotion processing and reinforcement.
-
The amygdala is particularly important in emotional learning and memory, particularly fear conditioning. It assesses emotional salience and generates appropriate emotional responses.
-
The hippocampus facilitates the formation of emotional memories, which are crucial for reinforcing future emotional responses based on past experiences.
-
The prefrontal cortex regulates emotions and decision-making processes, allowing for the modulation of emotional responses based on social norms and logical reasoning.
-
The nucleus accumbens plays a central role in the reinforcement circuit. It releases dopamine in response to reinforcing stimuli, reinforcing behaviors that lead to positive emotions.
Neurotransmitter Systems and Hormones
-
Dopamine pathways, particularly in the mesolimbic system, are associated with the reward circuit, reinforcing pleasurable emotions.
-
Serotonin influences mood, anxiety, and happiness, and its dysregulation is linked to mood disorders.
-
Norepinephrine affects arousal and alertness, mediating stress responses and emotional intensity.
-
Cortisol, the “stress hormone,” is released in response to stress and can reinforce the emotional response to stressful or threatening situations.
Neural Plasticity
Repeated emotional experiences can lead to neural plasticity, where the brain’s structure and function are modified, reinforcing specific emotional responses. This is evident in phenomena like fear conditioning and habituation (see images below).
Synaptic Strengthening
Synaptic strengthening, or long-term potentiation (LTP), is a process where repeated stimulation of a neural pathway increases the strength of synaptic transmission. This mechanism is crucial for learning and memory and applies to emotional learning as well. Emotional experiences can lead to LTP in neural circuits associated with emotion, reinforcing these emotional responses.
Neural Circuits and Feedback Loops
The brain operates through complex neural circuits and feedback loops that involve cortical and subcortical regions. The prefrontal cortex (PFC), which is involved in decision-making and moderating social behavior, can regulate the emotional responses generated by the limbic system. Through top-down regulation, the PFC can reinforce or inhibit emotional responses based on past experiences and cognitive appraisal of the situation.
Emotional Reinforcement
Emotions themselves can act as reinforcers. For example, pleasure from social interactions can reinforce social behaviors, while discomfort from fear can reinforce avoidance behaviors. Emotional reinforcement is integral to learning from our environment and experiences, shaping our future behaviors and emotional responses.
Neurocognitive Perspective
Neurobiological Basis of Emotions:
-
The neurobiology field has highlighted the role of specific brain regions, such as the amygdala and prefrontal cortex, in processing emotions. The amygdala, for instance, is crucial in fear and aggression, while the prefrontal cortex regulates these responses.
-
Neurotransmitters like dopamine, serotonin, and oxytocin play significant roles in mood regulation and emotional bonding. For example, oxytocin release during positive interactions with humans and dogs can reinforce the attachment.
Cognitive and Social Influences on Emotions:
-
Dogs have shown the ability to interpret human emotional signals, such as tone of voice and body language, which can influence their emotional states. For example, a dog might sense its owner’s stress and react accordingly, reinforcing its emotional response to human states.
-
Social learning is another key aspect. Dogs can learn from observing other dogs or humans, which can influence their emotional reactions. For instance, a dog might learn to associate a vet visit with fear if it observes other dogs exhibiting fearful behavior in that context.
Genetic and Temperamental Factors:
-
Genetics play a role in predisposing dogs to certain emotional responses. For instance, some breeds are more prone to anxiety or fearfulness, which can be reinforced through their interactions with their environment.
-
Individual temperament, influenced by genetics and early life experiences, also affects how a dog processes and reinforces emotional states.
Cognitive-Appraisal Theory:
-
This theory suggests that an individual’s perception and evaluation of an event influence their emotional experience. The cognitive appraisal process involves two stages: primary appraisal (assessing whether an event is positive, neutral, or negative) and secondary appraisal (considering one’s ability to cope with the event’s outcome). While more applicable to humans due to the complexity of cognitive processes involved, aspects of this theory can relate to animals in how they assess their environment and how this assessment influences their emotional state.
Learning Theories Perspective
-
Classical Conditioning: Pavlov demonstrated this learning process, which involves associating a neutral stimulus with an emotionally charged stimulus, leading to a conditioned emotional response. For example, if a sound (neutral stimulus) is repeatedly paired with a painful stimulus (unconditional stimulus), the sound alone can evoke a fear response (conditional response).
-
Operant Conditioning: B.F. Skinner’s theory posits that behaviors followed by positive outcomes are more likely to be repeated, while those followed by adverse outcomes are less likely. In the context of emotions, behaviors that result in positive emotional states (like happiness or relief) are reinforced. In contrast, those leading to negative emotions (like fear or sadness) might be avoided.
-
Social Learning Theory: Developed by Albert Bandura, social learning theory posits that learning occurs within a social context and can happen purely through observation or instruction, even without motor reproduction or direct reinforcement. This theory emphasizes the role of observational learning, imitation, and modeling in reinforcing emotions and behaviors. For instance, pets might observe and mimic the emotional responses of their human caretakers or other animals in their environment, reinforcing certain emotional behaviors without direct experience.
Behaviorism Perspective
Behaviorism, particularly as articulated by B.F. Skinner focuses on observable behaviors and the environmental factors that influence them. Mechanisms of reinforcement in behaviorism include:
-
Operant Conditioning (Reinforcers) involves reinforcing desired behaviors through adding (positive reinforcement) or removing a stimulus (negative reinforcement). Skinner’s experiments with rats and pigeons illustrated how reinforcement schedules (e.g., fixed-ratio, variable-ratio) affect behavior persistence.
-
Shaping: This technique involves reinforcing successive approximations of a desired behavior, gradually guiding an individual toward the target behavior through incremental reinforcement.
-
Behavior Modification: Techniques derived from behaviorism, such as token economies and reinforcement schedules, are widely used in educational and therapeutic settings to promote desired behaviors.
In the context of behaviorism, I want to mention active and vicarious learning in shaping and reinforcing emotional responses. Active learning involves direct engagement with experiences that evoke emotions, such as through expressive arts or role-playing. This approach allows individuals to confront and express their feelings, reflect on their emotional experiences, and receive feedback, thereby enhancing emotional awareness and resilience. Conversely, vicarious learning involves learning about emotions through observing others. By witnessing how others handle emotional situations—through stories, films, or real-life interactions—individuals gain insights into managing their emotions and internalizing social norms regarding emotional expression. This type of learning also fosters empathy, helping individuals connect more deeply with others.
Expanding on the Behaviorist Approach
-
Detailed Mechanisms of Classical Conditioning: Classical conditioning in dogs often involves creating associations between stimuli and emotional responses. For example, if a dog repeatedly hears a specific sound (like a whistle) before mealtime, it may begin to associate that sound with the pleasure of eating, leading to excitement upon hearing the whistle.
-
Operant Conditioning Nuances: In operant conditioning, emotional reinforcement is closely tied to the consequences of the dog’s behavior. If a behavior leads to a positive outcome (like receiving a treat or affection), the dog will likely repeat the behavior, reinforcing the associated positive emotion.
-
Negative reinforcement also plays a role, where a behavior that leads to the cessation of an unpleasant stimulus (like stopping a loud noise) reinforces the behavior through relief or decreased anxiety.
-
Emotional Extinction and Counterconditioning: Extinction occurs when a conditioned emotional response diminishes over time as the conditional stimulus is presented without the unconditional stimulus. For example, if the sound of a doorbell is no longer followed by visitors, a dog’s excitement or anxiety may gradually decrease. Counterconditioning involves replacing an unwanted emotional response with a more desirable one. This is often used in behavior modification programs to address fearful or aggressive behaviors.
Ethology Perspective
Ethology, the study of animal behavior in natural environments, provides insights into the evolutionary significance of reinforcement. Mechanisms include:
-
Natural Selection: Evolutionary processes reinforce behaviors that enhance survival and reproductive success.
-
Innate Behaviors and Learning: Ethologists study how certain behaviors are instinctual while others are learned through reinforcement.
-
Social Learning: Ethology also examines how animals learn from observing others, with reinforcement playing a role in adopting behaviors within social groups. This can include learning through imitation or the reinforcement of social hierarchies.
Ethology also offers a robust framework for comprehending how behaviors are shaped by their associated costs and benefits. As a discipline focused on the study of animal behavior, ethology underscores the necessity of assessing the outcomes of actions through the perspective of net value.
A central concept within ethology is the Net Value of Behavior (BNV), which is defined as the difference between the benefits (B) and the costs (C) associated with a behavior. This relationship is mathematically expressed by the formula: BNV = B – C. A positive BNV signifies that the benefits exceed the costs, thereby making the behavior advantageous. Conversely, a negative BNV indicates that the costs surpass the benefits, rendering the behavior less favorable. When BNV is zero, the behavior is considered neutral, presenting no distinct advantage or disadvantage.
The frequency of a behavior (BF) is closely correlated with its net value, as represented by the equation BF = f(BNV). This implies that the probability of a behavior being repeated is directly proportional to its net value. For example, if an animal encounters a behavior that provides substantial benefits with minimal costs, it is more likely to re-engage in that behavior, thereby reinforcing the positive emotional response associated with it.
Merging Neuroscience and Behaviorism
-
Emotional Learning: Both fields acknowledge that emotional responses can be learned and modified based on experience (fig. 3, 4 and 5). Neuroscience explains the underlying brain mechanisms, while behaviorism focuses on observable behaviors and their consequences.
-
Feedback Loops: Behaviorism’s reinforcement principles align with the neuroscientific understanding of reward pathways. Behaviors leading to positive emotional states activate reward circuits in the brain, possibly reinforcing these behaviors.
-
Adaptive Significance: Emotions have an adaptive role from an evolutionary perspective. Neuroscience explains this through the brain’s mechanisms for survival (e.g., fear response), while behaviorism interprets it through the lens of behaviors that have been reinforced over time for survival advantages.
-
Affective Neuroscience: Affective neuroscience explores the neural mechanisms of emotions, offering insights into how emotions are processed and reinforced at the biological level. This field studies how different brain regions (like the amygdala, prefrontal cortex, and hippocampus) and neurotransmitter systems contribute to the formation and reinforcement of emotional experiences.
-
Emotional Regulation Theory: Emotional regulation involves the processes by which individuals influence which emotions they have, when they have them, and how they experience and express these emotions. This theory is complex and primarily human-focused, but aspects can be applied to understanding how pets manage or modify their emotional responses based on their environment or training. For example, pets can learn to regulate their emotional responses to stimuli (like reducing fearfulness or aggressiveness) through training and socialization.
-
Attachment Theory: Originally formulated to describe the dynamics of human relationships, attachment theory has also been applied to understand the bonds between humans and pets. This theory suggests that the quality of attachment affects emotional security and behavior. Secure attachments can reinforce positive emotions and adaptive behaviors, while insecure attachments may lead to anxiety and maladaptive behaviors. Pets, like humans, can exhibit signs of secure or insecure attachment based on their interactions with their caregivers, influencing their emotional well-being. however, the tiny line of anthropomorphism should be critically evaluated.
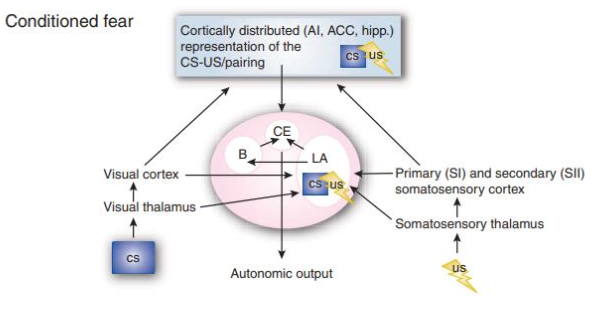
Fig. 3. A neural model of non-social fear learning in humans. Connectivity is bidirectional, although the arrows point in one direction and describe the flow of information between different brain functional regions. Fear conditioning occurs by associating the conditional stimulus (CS) representation with the somatosensory representation of the unconditional stimulus (US). The lateral nucleus (LA), where CS and US converge, is the learning site. The amygdala receives information from the hippocampal memory system (Hipp), anterior insula (Al), and anterior cingulate cortex (ACC), which contains secondary representations of the CS and US (information about the learning context and the internal state of the organism. From Olson et al. (2007).
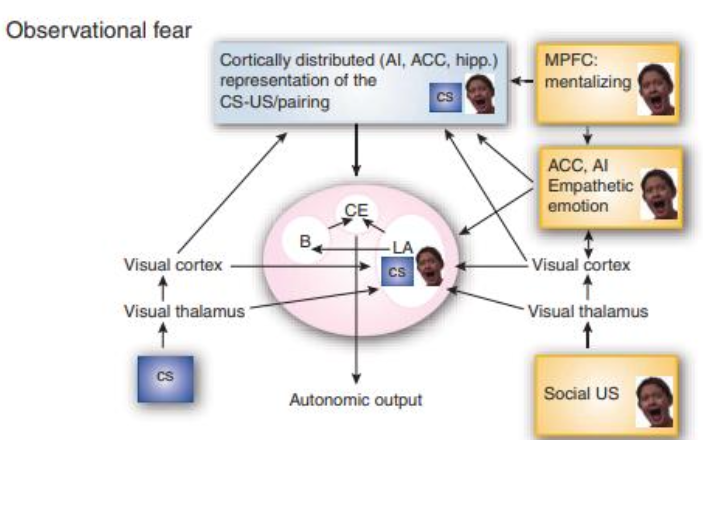
Fig. 4. In observational fear learning, the CS is modified by association with a representation of the distressed peer, serving as US. The hypothesis of CS and US representations converge in the LA. The strength of the US can be modified by MPFC input related to the mental interpretation of the other and the cortical representation of empathic pain through the ACC and the LA. From Olson et al. (2007).
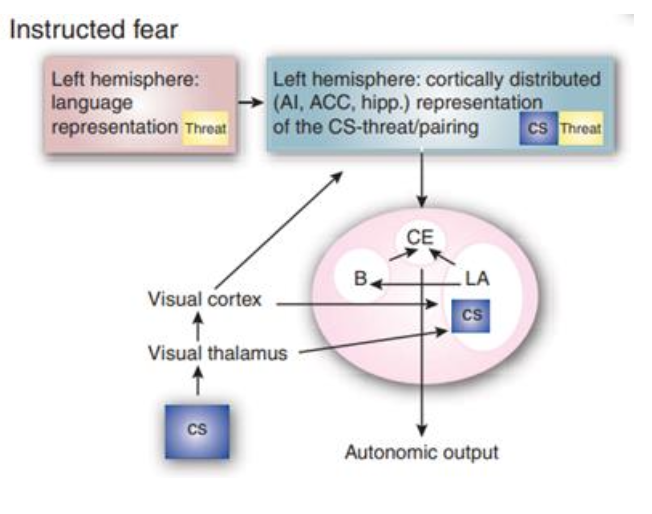
Fig. 5. Fear-instructed learning occurs by modifying the processing of the visual representation of the CS through an abstract representation of the threat. Consequently, contingency is likely to rely heavily on the hippocampal memory system rather than being encoded in the amygdala. From Olson et al. (2007).
Integrated Emotional Model (IEM)
My suggestion on this topic introduces the Integrated Emotional Model (IEM) for animal training, a direct approach that emphasizes animals’ emotional, cognitive, and social complexities. This suggestion encourages a nuanced understanding of animal behavior, steering clear of anthropomorphism—the tendency to attribute human emotions and motivations to animals, leading to misunderstandings and misinterpretations of their actions and needs.
The IEM focuses on accurately recognizing and interpreting animals’ behavioral signals, a crucial skill for tailoring training to each animal’s unique circumstances and promoting their well-being. By carefully avoiding the pitfalls of anthropomorphism and grounding our practices in evidence-based understanding of animal behavior, the IEM offers a realistic, respectful framework for enhancing animal training, ensuring it is both effective and considerate of the animal’s emotional state.
In practice, the IEM challenges trainers and owners to be more attuned to the subtle nuances of animal behavior. It underscores the importance of creating training scenarios that are physically safe and emotionally supportive, allowing animals to learn and thrive without fear or distress.
The Integrated Emotional Model represents a forward-thinking approach to training that prioritizes the health and welfare of animals, paving the way for more humane and educated interactions between humans and the animals in their care.
Intrinsic Emotional Factors:
-
An animal’s emotional responses are not solely products of conditioning but are also influenced by inherent factors like genetics, neurobiology, and individual temperament.
-
Neuroscientific research, including fMRI studies, indicates that dogs have complex neural networks for processing emotions, implying an innate aspect of their emotional experiences.
-
Understand and accommodate the living (biotic) elements, such as social interactions, health, and nutrition, alongside non-living (abiotic) factors like temperature, habitat structure, and sensory stimuli. This balance ensures training is adapted to each animal’s environmental context, promoting comfort, focus, and optimal learning conditions.
Cognitive and Social Elements:
-
Dogs demonstrate significant cognitive abilities, including understanding human signals and influencing their emotional responses beyond simple conditioning.
-
Studies in canine social cognition illustrate that dogs are sensitive to human emotions, indicating that their emotional states are shaped by their interactions with humans and their environment, not just through reinforcement. Incorporating a critical perspective on anthropomorphism, it’s imperative to balance our beliefs with scientific understanding. While recognizing the emotional lives of the animals, we must avoid projecting human emotions and motivations onto them, which could lead to misinterpretations and inappropriate responses to their needs. This critical stance encourages a respectful and nuanced approach to interpreting animals’ behaviors and emotional states.
Neurobiological Insights:
-
The exploration of neurobiology has enriched our understanding of animal emotions, identifying specific brain regions and neurotransmitter systems involved in emotion processing. This parallel with human emotional neurobiology offers insights into the evolutionary continuity of emotional mechanisms across species, emphasizing the importance of a neurobiologically informed approach to training and health care.
-
The role of neuroendocrine factors, like the oxytocin system, in mediating social bonding and stress responses highlights the biochemical underpinnings of animal emotions. These insights underscore the importance of considering how emotional well-being is linked to physical health and advocating for training and care practices that support both aspects.
Critical Thinking Models for Practical Analysis:
Systemic Thinking:
-
Systemic thinking encourages trainers to view the training process as part of a larger system, recognizing the interconnectedness of various elements affecting an animal’s learning and behavior:
-
Identify Interconnections: Understand how different aspects of the animal’s life, such as home environment, interactions with humans and other animals, and previous training experiences, influence their behavior and response to training.
-
Holistic Approach: Develop training plans considering these interconnections, aiming for consistency across different environments and situations. This approach ensures that behaviors learned in training are transferable and stable across contexts.
Feedback Loops:
-
Pay attention to feedback from the animal during training sessions. This includes direct responses to signals and more subtle signs of stress, confusion, or engagement. Adjust training strategies based on this feedback to create a more responsive and adaptive training process.
Critical Thinking:
-
Critical Thinking in animal training involves questioning assumptions, analyzing information, and evaluating the effectiveness of training methods. Practical steps include:
-
Question Assumptions: Challenge common beliefs about animal behavior and training methods. This could involve re-evaluating the use of certain reinforcers or punishments based on the latest research or the specific needs of the animal.
-
Evidence-Based Decisions: Use scientific research and data to inform training practices. This means staying updated on the latest animal behavior and neurobiology studies and applying those insights to training methodologies.
Reflect and Evaluate:
-
Regularly assess the outcomes of training sessions, not just in terms of immediate behavior changes but also considering long-term effects on the animal’s emotional and physical well-being. This reflective practice allows for continual improvement and adaptation of training methods.
The “Comfort” Discussion
To finish, I want to address this popular trend. The notion that emotions are immutable has been popularized, in part, by examples of comforting dogs in distress. This belief, appealing due to its emotional resonance and the desire to alleviate animal suffering, ironically can contribute to its proliferation. This trend continues despite contradicting the academic consensus, as shown in the documents and references below, and is propelled by social reinforcement and cultural survival mechanisms.
Many pet owners and trainers advocate the practice of comforting dogs during storms despite its contentious nature and extensive documentation in academic circles. Exploring neuroscience, learning theory, and canine behavior sheds light on the potential drawbacks of this approach. However, this discussion raises several challenging questions that warrant further exploration:
-
How do people define comfort, and how do they apply it in various contexts? The subjective nature of comfort can vary significantly among individuals and cultures. What one person perceives as comforting may be distressing to another. This variability complicates the understanding of emotional responses and the appropriateness of comforting behaviors.
-
What are the long-term effects of comforting behaviors on human and animal emotional development? While immediate comfort may alleviate distress, it is essential to consider whether such interventions promote healthy emotional regulation or inadvertently reinforce maladaptive behaviors. For instance, does comforting a dog during a storm help it cope with future storms, or does it create a dependency on human reassurance?
-
How do societal norms and cultural beliefs shape our understanding of emotional expression and comfort? Different cultures have varying beliefs about emotional expression, which can influence how comfort is perceived and enacted. Understanding these cultural dimensions can provide insight into the broader implications of comforting behaviors and their acceptance within different communities.
-
What role does individual temperament play in the effectiveness of comforting strategies? Just as humans have different emotional responses based on personality traits, dogs may also respond differently to comforting behaviors. Some dogs may thrive on reassurance, while others may become more anxious. How can we tailor our approaches to accommodate these individual differences?
-
How do advancements in neuroscience inform our understanding of emotional responses in humans and animals? As neuroscience research continues to evolve, it offers new insights into the mechanisms underlying emotional experiences. How can this knowledge be applied to improve our approaches to comforting behaviors, ensuring they are grounded in scientific understanding rather than anecdotal evidence?
-
What ethical considerations arise from the practice of comforting animals in distress? The desire to alleviate suffering is noble, but it is crucial to examine whether our interventions are genuinely beneficial for the animal’s emotional well-being. Are we prioritizing our emotional needs over the animal’s natural coping mechanisms?
Many pet owners and trainers advocate the practice of comforting dogs during storms despite its contentious nature and extensive documentation in academic circles. Exploring neuroscience, learning theory, and canine behavior sheds light on the potential drawbacks of this approach:
-
Emotional Reinforcement, A Scientific Fact: Contrary to popular belief, neuroscience supports the concept that emotions can be reinforced, with experiences shaping or modifying the brain’s neural pathways, thereby influencing emotional responses. This well-established scientific principle has yet to fully permeate the dog training industry, where misconceptions still persist.
-
The “Double-Edged Sword” of Comfort: Fear conditioning illustrates that emotions and behaviors associated with fear can be reinforced. The effectiveness of comfort as reinforcement varies by individual and situation. If a dog interprets comfort as a positive reinforcement during a storm, this could unintentionally encourage fearful behavior. Alternatively, if the comfort alleviates fear, it may serve as negative reinforcement, encouraging the dog to seek comfort in future stressful situations.
-
The Importance of Fostering Independence: Over-reliance on comfort can hinder a dog’s development of independent coping mechanisms. Encouraging autonomy while providing safe spaces is crucial for a dog’s emotional health, especially in the absence of their owner.
-
The Impact of the Owner’s Emotional State: Dogs are highly sensitive to human emotions, and an owner’s distress can amplify a dog’s fear, creating a compounded reaction to both the storm and the owner’s state of mind.
-
Inconsistency and Confusion: The unpredictable nature of storms, coupled with inconsistent human reactions, can result in confusing and potentially detrimental comforting practices, affecting the dog negatively.
-
Towards a More Informed Approach: Although the intention behind comforting dogs is compassionate, a deeper understanding of canine emotions and behavior underscores the necessity of a more informed approach. Overlooking scientific research and the individual needs of each dog, as well as failing to consider the specific context, can perpetuate the notion of dogs as perpetually vulnerable. This “poor dog syndrome” becomes a marketing niche, detracting from the need to provide accurate information and promote resilience.
Ethological models also might provide a differentiated approach to understanding behavior by evaluating its net value—benefits minus costs—based on observable actions, as mentioned above (Ethology Perspective). This framework is particularly useful for analyzing behaviors like a dog’s response to storms and the subsequent comforting actions of an owner. For instance, when a storm induces fear in a dog, the owner’s comforting behavior can be assessed by quantifying observable benefits, such as physical comfort and perceived safety, against observable costs, such as the time spent and potential disruption to the owner’s routine. A positive net value indicates that the benefits of comforting the dog outweigh the costs, suggesting that this behavior will likely be repeated. Conversely, the behavior may become less frequent if the net value is neutral or negative, meaning the costs outweigh the benefits. Although ethology effectively captures behavioral patterns’ dynamics and consequences, it does not measure emotional experiences directly. Instead, it relies on observable actions and outcomes to infer the relative value of behaviors and their impact on future actions.
From a psychological perspective, the owner’s behavior can be understood through concepts such as attachment and emotional regulation. The owner feels a compelling urge to engage in comforting behaviors, driven by a desire to alleviate the dog’s visible distress. This need for comfort may stem from the owner’s own emotional discomfort in witnessing the dog’s fear, leading them to seek a sense of control and purpose in the situation.
It is important to note that in some cases, the owner benefits more from these interactions than the dog. For instance, when the owner comforts the dog during a storm, they might experience an emotional boost from alleviating their own feelings of helplessness or anxiety. This act of comforting can give the owner a sense of purpose and control over a stressful situation, potentially more than it alleviates the dog’s fear. This perspective shifts the focus from the dog’s needs to the owner’s psychological reinforcement.
I could use the term “empathy” here, but I won’t. The term “empathy” is often used in discussions about dog training and behavior, sometimes as a marketing trend. Although empathy suggests a deep understanding of another’s feelings, it can be misleading in the context of dog behavior. Owners may assume that their comforting actions are what the dog truly needs, but this assumption can overlook their individual coping mechanisms and emotional needs. This reliance on a superfluous concept of empathy in dog training can unintentionally neglect the dog’s ability to develop resilience. Instead of fostering independence, the owner’s comforting behavior may inadvertently reinforce the dog’s fearful behavior, creating a cycle of dependency. The owner, believing they are acting empathetically, may not recognize that their actions could hinder the dog’s emotional growth and ability to cope with stressors independently.
Additionally, the owner’s anxiety and need for reassurance can have a significant influence. In the face of a storm, the owner may experience heightened feelings of vulnerability and uncertainty. By comforting the dog, they channel their anxiety into nurturing actions, creating a temporary sense of control over the chaotic environment. This dynamic can lead to a cycle where the owner’s need for comfort becomes a coping mechanism, allowing them to manage their emotional discomfort while potentially neglecting the dog’s need to learn self-soothing strategies.
This situation can raise ethical questions about dependency and autonomy. The dog’s reliance on his owner for comfort during storms may foster a sense of dependency that complicates his ability to cope with fear independently. The owner’s well-intentioned actions, while rooted in concern, may inadvertently hinder the dog’s emotional growth and resilience. Seeking comfort or help is a common coping mechanism in response to fear. When an animal feels threatened or anxious, it often seeks proximity to a trusted figure as part of its fear response (e.g., Hennessy & Kaiser, 2006). This behavior is an aspect of social buffering, a well-documented phenomenon where social support mitigates stress responses in fearful situations (Hostinar & Gunnar, 2013). This paradox highlights the tension between the desire to protect and the necessity of allowing space for self-soothing and personal development.
Furthermore, the concept of existentialism comes into play as both the dog and the owner navigate their experiences of fear and anxiety. The dog’s fear of storms represents a fundamental aspect of existence—the confrontation with the unknown and the uncontrollable. The owner’s comforting behavior, while an attempt to mitigate this fear, also reflects their struggle with the unpredictability of life. In seeking to provide comfort, the owner grapples with their existential concerns, revealing the interconnectedness of their emotional states.
In conclusion of this topic, to provide comfort to dogs during storms, we need to maintain a balance between compassion and scientific understanding. Without the reinforcement or inhibition of emotions, survival and social relationships would be profoundly compromised, as emotions serve as fundamental guides for adaptation. Fear, for instance, mobilizes responses that protect against imminent dangers, while joy strengthens social bonds and behaviors that benefit group living. Without this emotional mechanism, an individual would neither learn to avoid threats nor seek experiences that promote safety and well-being, resulting in random and uncoordinated behaviors.
In discussions about comforting dogs during thunderstorms, the debate often drifts into moralistic territory, disregarding the function of emotions. Emotions are not simply arbitrary feelings to be indulged or suppressed; they are integral to the survival and social cohesion of both individuals and groups. In relationships, the absence of emotional reinforcement would erode trust and empathy, leaving bonds fragile and incapable of supporting the complex interactions required for coexistence and cooperation.
It is also important to consider the complicated emotional states of canines, the tendency for subjective assumptions, the tiny line of anthropomorphism, and the impact of human behavior on them. This will help dogs develop the skills they need to tackle challenges independently. We should move past simplistic notions of vulnerability and strive to develop a more resilient and informed relationship between dogs and their owners. Neuroscience supports the concept that emotions can be reinforced, indicating that comforting a dog during a storm might unintentionally reinforce fearful behavior. Whether comfort acts as reinforcement depends on the individual and the situation; not all individuals will perceive comfort similarly, and not all situations will lead to comfort acting as a reinforcer.
Multimedia
- Playlist “Animal Like Us”: https://www.youtube.com/playlist?list=PLTOUyQcU3JG38kwqjUnxhn0zFRHA5dB09
- https://youtu.be/a9LjXHtLvlY
Links
-
https://www.sciencedirect.com/science/article/pii/S0168159118300285
-
https://www.researchgate.net/publication/356284307_Animal_Emotions-Do_Animals_Feel_as_We_Do
-
https://www.facebook.com/tilde.d.jensen/videos/4130290023688258/?idorvanity=318134942711314
-
https://www.researchgate.net/publication/362677075_Neurophysiology_of_Emotions
-
https://www.researchgate.net/publication/319269659_The_Influences_of_Emotion_on_Learning_and_Memory
-
https://www.emory.edu/LIVING_LINKS/publications/articles/deWaal_2011b.pdf
References
Adolphs R. (2013). The biology of fear. Current biology: C.B., 23(2), R79–R93. https://doi.org/10.1016/j.cub.2012.11.055
Adolphs, R., & Anderson, D. J. (2018). The neuroscience of emotion: A New Synthesis. Princeton University Press.
Barata, R. (2012). Dominance in Dogs: A Comprehensive Scientific Compilation. Human-Animal Science.
Barrett, L. F., & Satpute, A. B. (2013). “Large-scale brain networks in affective and social neuroscience: Towards an integrative functional architecture of the brain.” Current Opinion in Neurobiology, 23(3), 361-372.
Beck, J. S. (2011). Cognitive behavior therapy: Basics and beyond. Second Edition. Guilford Press.
Bekoff, M. (2007). The emotional lives of animals: A leading scientist explores animal joy, sorrow, and empathy—and why they matter. New World Library.
Bradshaw, J. (2011). Dog sense: How the new science of dog behavior can make you a better friend to your pet. Basic Books.
Cannon, W. B. (1927). The James-Lange Theory of Emotions: A Critical Examination and an Alternative Theory. Psychological Review, 34(6), 106.
Choudhury, S., & Slaby, J. (2011). Critical Neuroscience: A Handbook of the Social and Cultural Contexts of Neuroscience. Wiley-Blackwell.
Dalgleish, T. (2004). The emotional brain. Nature Reviews Neuroscience, 5(7), 583–589. https://doi.org/10.1038/nrn1432
Damasio, A. R. (1994). Descartes’ Error: Emotion, Reason, and the Human Brain. Putnam.
Damasio, A. (1999). The Feeling of What Happens: Body and Emotion in the Making of Consciousness. Harcourt.
Della Sala, D. (Ed.). (2020). Encyclopedia of behavioral neuroscience (2nd ed.). Elsevier.
Dinsmoor, J. A. (2004). The etymology of basic concepts in the experimental analysis of behavior. Journal of the Experimental Analysis of Behavior, 82(3), 311–316. https://doi.org/10.1901/jeab.2004.82-311
Ekman P. (1992). Are there basic emotions?. Psychological review, 99(3), 550–553. https://doi.org/10.1037/0033-295x.99.3.550
Ekman, P. (1994). All emotions are basic. In P. Ekman & R.J. Davidson (Eds.), The nature of emotion: Fundamental questions (pp. 15–19). New York: Oxford University Press.
English, H. B., & English, A. C. (1958). A comprehensive dictionary of psychological and psychoanalytical terms: A guide to usage. Longmans, Green.
Farah, M. J. (2011). Neuroscience and neuroethics in the 21st century. In Oxford University Press eBooks. https://doi.org/10.1093/oxfordhb/9780199570706.013.0172
Fitzpatrick, S. (1990). Russians on the psyche: A review. Science, 248, 881-883.
Gallistel, C. R., Fairhurst, S., & Balsam, P. (2004b). The learning curve: Implications of a quantitative analysis. Proceedings of the National Academy of Sciences, 101(36), 13124–13131. https://doi.org/10.1073/pnas.0404965101
Goldman, J. G. (2012). What is classical conditioning? (And why does it matter?). Scientific American. https://www.scientificamerican.com/blog/thoughtful-animal/what-is-classical-conditioning-and-why-does-it-matter/
Gordon, R. M. (2008). Beyond mindreading. Philosophical Explorations, 11(3), 219–222. https://doi.org/10.1080/13869790802239250
Hennenlotter, A., Dresel, C., Castrop, F., Ceballos-Baumann, A.O., Wohlschläger, A.M., & Haslinger, B. (2009). The link between facial feedback and neural activity within central circuitries of emotion–new insights from botulinum toxin-induced denervation of frown muscles. Cerebral cortex, 19 3, 537-42 .
Hennessy, M. B., & Kaiser, S. (2006). Social buffering of the stress response: Diversity, mechanisms, and functions. Frontiers in Neuroendocrinology, 27(3), 274-286. https://doi.org/10.1016/j.yfrne.2009.06.001
Hochschild, A. R. (1983). The managed heart: Commercialization of human feeling. University of California Press.
Hostinar, C. E., & Gunnar, M. R. (2013). The developmental psychobiology of stress and emotion in childhood. In R. M. Lerner, M. A. Easterbrooks, J. Mistry, & I. B. Weiner (Eds.), Handbook of psychology: Developmental psychology (2nd ed., pp. 121–141). John Wiley & Sons, Inc..
James, W. (1884). What is an Emotion?. Mind, 9(34), 188-205.
Jardat, P., & Lansade, L. (2021). Cognition and the human-animal relationship: a review of the sociocognitive skills of domestic mammals toward humans. Animal Cognition, 25(2), 369–384. https://doi.org/10.1007/s10071-021-01557-6
Kolb, B., & Gibb, R. (2014). Brain Plasticity and Behavior. Annual Review of Psychology, 65, 441-470.
LeDoux, J. E. (1996). The emotional brain: The mysterious underpinnings of emotional life. Simon & Schuster.
LeDoux, J. E. (2012). Rethinking the Emotional Brain. Neuron, 73(4), 653-676.
Mackintosh, N. J. (1974). The psychology of animal learning. Academic Press.
MacLean, P. D. (1990). The Triune Brain in Evolution: Role in Paleocerebral Functions. Plenum Press.
Mendl, M., Burman, O. H., & Paul, E. S. (2010). An integrative and functional framework for the study of animal emotion and mood. Proceedings of the Royal Society B: Biological Sciences, 277(1696), 2895-2904.
Merleau-Ponty, M. (1962). Phenomenology of perception. Routledge.
Miklosi, A. (2007). Dog Behaviour, Evolution, and Cognition. Oxford University Press.
Morris, R. W., & Bouton, M. E. (2006). Effect of unconditioned stimulus magnitude on the emergence of conditioned responding. Journal of Experimental Psychology: Animal Behavior Processes, 32(4), 371–385. https://doi.org/10.1037/0097-7403.32.4.371
Neisser, U. (1976). Cognition and reality: Principles and implications of cognitive psychology. Freeman.
Neimeyer, R. A. (1995). An invitation to constructivist psychotherapies. In R. A. Neimeyer & M.J. Mahoney (Eds.), Constructivism is psychology (pp. 1–8). Washington, DC: American Psychological Association.
Olsson, A., & Phelps, E. A. (2007). Social learning of fear. Nature Neuroscience, 10(7), 1095–1102. https://doi.org/10.1038/nn1968
Olson, M. & Hergenhahn, B. R. (2016). An Introduction to Theories of Learning, Ninth edition. Psychology Press.
Panksepp, J. (1994). The basics of basic emotion. In P. Ekman & R.J. Davidson (Eds.), The nature of emotion: Fundamental questions (pp. 20–24). New York: Oxford University Press.
Panksepp, J. (1998). Affective Neuroscience: The Foundations of Human and Animal Emotions. New York, NY: Oxford University Press.
Panksepp, J. (2005). Affective consciousness: Core emotional feelings in animals and humans. Consciousness and Cognition, 14(1), 30-80.
Panksepp, J., & Biven, L. (2012). The Archaeology of Mind: Neuroevolutionary Origins of Human Emotions. Norton & Company.
Pavlov, I. P. (1927). Conditioned Reflexes: An Investigation of the Physiological Activity of the Cerebral Cortex. Oxford University Press.
Pavlov, I. P. (1960). Conditioned reflexes: An investigation of the physiological activity of the cerebral cortex (G. V. Anrep, Trans.). Dover. (Original work published 1927).
Phelps, E. A., & LeDoux, J. E. (2005). Contributions of the amygdala to emotion processing: From animal models to human behavior. Neuron, 48(2), 175-187.
Ray, W. J. (2024). Fundamentals of brain and behavior: An Introduction to Human Neuroscience.
Reevy, G., Ozer, Y. M., & Ito, Y. (2010). Encyclopedia of Emotion. Abc-clio.
Rescorla, R., & Wagner, A. R. (1972). A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement.
Rolls, E. T. (2013). Emotion and Decision Making Explained. Oxford University Press.
Roskies, A. L., & Morse, S. J. (2013). Neuroscience and the law. In Oxford University Press eBooks (pp. 240–255). https://doi.org/10.1093/acprof:oso/9780199859177.003.0009
Sartre, J. P. (1943). Being and Nothingness. Washington, DC: Washington Square Press.
Schacter, S., & Singer, J. E. (1962). Cognitive, social, and physiological determinants of emotional state. Psychological Review, 69(5), 379-399. https://doi.org/10.1037/h0046234
Schoenfeld, W. N. (1978). “Reinforcement” in behavior theory. Pavlovian Journal of Biological Science, 13, 135–144. https://doi.org/10.1007/BF03001385
Schoenfeld, W. N. (1995). “Reinforcement” in behavior theory. The Behavior Analyst, 18(1), 173–185. https://doi.org/10.1007/bf03392705
Shepherd, S. V. (2017). The Wiley Handbook of Evolutionary Neuroscience. John Wiley & Sons.
Shettleworth, S. J. (2010). Cognition, Evolution, and Behavior. Oxford University Press.
Skinner, B. F. (1953). Science and Human Behavior. Macmillan.
Sperber, D., & Wilson, D. (1995). Relevance: Communication and Cognition. Blackwell Publishing.
Sterelny, K. (2003). Thought in a Hostile World: The Evolution of Human Cognition. Blackwell Publishing.
Wallis, L., Range, F., Müller, C., Serisier, S., Huber, L., & Viranyi, Z. (2013). Age effects on interspecific communicative abilities of domestic dogs. Journal of Veterinary Behavior, 8(4), e31. https://doi.org/10.1016/j.jveb.2013.04.016
Windholz, G. (1989). The discovery of the principles of reinforcement, extinction, generalization, and differentiation of conditional reflexes in Pavlov’s laboratories. The Pavlovian Journal of Biological Science, 24(2), 35–42. https://doi.org/10.1007/bf02964534
Yokoyama, O. (2023). Pavlov on the Conditional Reflex. Oxford University Press eBooks. https://doi.org/10.1093/oso/9780190941871.001.0001